Une association entre la structure génétique des bactéries intestinales et la cognition, ainsi que les affections neuroanatomiques humaines, a été établie.
Le corps humain héberge une multitude de micro-organismes, notamment des bactéries, des champignons, des virus, et d’autres entités biologiques, constituant ainsi le microbiote de l’organisme. Les populations prédominantes de ces micro-organismes sont situées principalement dans l’intestin, formant ainsi le microbiote intestinal. En raison de l’impact significatif de ce microbiote sur la santé et le bien-être humains, les chercheurs affiliés à la Harvard Medical School ont entrepris d’investiguer la présence d’un lien génétique entre les bactéries intestinales et certains comportements humains.
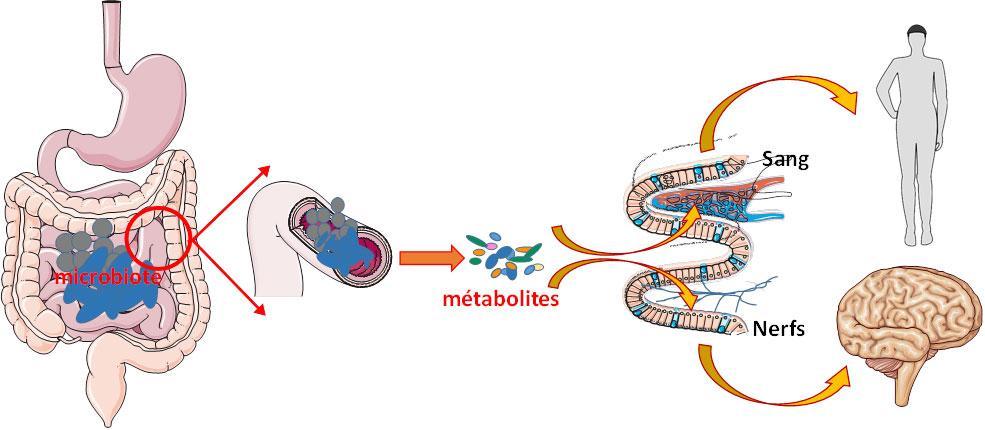
Les composés métaboliques générés par le microbiote intestinal peuvent être acheminés vers la circulation sanguine par les cellules de la paroi intestinale. Ces métabolites ont la capacité d’interagir avec l’ensemble des organes, y compris le cerveau, par le biais de la circulation sanguine. Le tractus gastro-intestinal est richement innervé de nerfs portant des récepteurs qui identifient diverses molécules, certaines d’entre elles étant produites par les bactéries intestinales. Ainsi, les métabolites issus de ces bactéries peuvent établir une interaction directe avec le cerveau via les connexions nerveuses.
INTRODUCTION
La relation étroite entre le système gastro-intestinal et le cerveau est évidente. Les signaux provenant du cerveau atteignent le tractus gastro-intestinal par le biais du système nerveux autonome et du système endocrinien, tandis que le tractus gastro-intestinal peut communiquer avec le cerveau par le nerf vague ainsi que par des molécules de signalisation endocriniennes et immunitaires (cytokines). De plus, les produits du métabolisme microbien générés dans le tractus gastro-intestinal peuvent influencer le cerveau, à la fois de manière indirecte en stimulant les systèmes nerveux entérique et immunitaire, et directe à travers des molécules qui pénètrent dans la circulation et traversent la barrière hémato-encéphalique. Des liens de causalité entre le microbiome intestinal et le développement neural, en particulier le développement atypique, sont de plus en plus identifiés. À la fois l’épidémiologie humaine et les modèles animaux pointent vers les effets des microbes intestinaux sur le développement du trouble du spectre de l’autisme et des taxons microbiens spécifiques ont été associés à la dépression et à la maladie d’Alzheimer. Cependant, des informations sur cet « axe microbiome-intestin-cerveau » dans le développement neurocognitif normal font défaut, en particulier au début de la vie.
Les premières années de la vie constituent des fenêtres de développement critiques tant pour le microbiome que pour le cerveau. On pense que le développement fœtal se produit dans un environnement stérile, mais les nouveau-nés sont rapidement colonisés à la naissance par contact avec le canal de naissance (en cas d’accouchement par voie basse), les soignants, la nourriture (lait maternel ou formule), et d’autres sources environnementales. Le microbiome précoce se caractérise par une faible diversité microbienne, une succession rapide et une évolution, et est dominé par les Actinobactéries, en particulier le genre Bifidobacterium, les Bactéroïdètes, notamment Bacteroides, et les Protéobactéries. Beaucoup de ces bactéries ont des capacités métaboliques spécialisées pour la digestion du lait maternel humain, telles que Bifidobacterium longum subsp. infantis et Bacteroides fragilis. Avec l’introduction des aliments solides, le microbiome intestinal subit une autre transformation catégorielle ; sa diversité augmente, et la plupart des taxons du microbiome infantile sont remplacés par des taxons plus semblables à ceux des microbiomes adultes. Les études antérieures se sont généralement concentrées soit sur les microbiomes infantiles, soit sur les microbiomes adultes, car réaliser des analyses statistiques à travers cette transition pose des défis particuliers. Néanmoins, puisque cette transition coïncide avec des fenêtres de développement neural critiques et des processus neurodéveloppementaux associés tels que la myélinisation, la neurogenèse et l’élagage synaptique, une investigation à travers cette frontière des aliments solides est importante.
Le cerveau d’un enfant subit des changements anatomiques, microstructuraux, organisationnels et fonctionnels remarquables au cours des premières années de la vie. À l’âge de 5 ans, le cerveau d’un enfant atteint plus de 85% de sa taille adulte et a atteint des niveaux de myélinisation proches de ceux d’un adulte, et le schéma des connexions axonales a été établi. Une grande partie de ce développement se produit dans des fenêtres discrètes appelées périodes sensibles (PS) au cours desquelles la plasticité neuronale est particulièrement élevée. Des preuves émergentes suggèrent que le timing et la durée des PS peuvent être en partie influencés par des signaux provenant du microbiome intestinal en développement. Ainsi, comprendre le spectre normal du développement sain du microbiome et sa relation avec le développement neurocognitif normal peut offrir des opportunités pour identifier plus tôt des développements atypiques et proposer des possibilités d’intervention.
Pour commencer à répondre à ce besoin, nous avons étudié la relation entre le microbiome intestinal et le développement neurocognitif dans une grande cohorte d’enfants en bonne santé et se développant de manière neurotypique depuis la petite enfance jusqu’à l’âge de 10 ans. Les communautés microbiennes intestinales ont été évaluées par séquençage métagénomique shotgun, permettant un profilage tant au niveau taxonomique que fonctionnel des gènes. Les compétences et capacités cognitives ont été mesurées à l’aide d’évaluations psychométriques appropriées à l’âge de la fonction cognitive, à savoir les Mullen Scales of Early Learning (MSEL), la Wechsler Preschool and Primary Scale of Intelligence, 4th Edition (WPPSI-4) et la Wechsler Intelligence Scale for Children, 5th Edition (WISC-V). Enfin, nous avons également évalué la structure cérébrale émergente à l’aide de l’imagerie par résonance magnétique (IRM). À travers une combinaison d’analyses statistiques classiques et d’apprentissage automatique (ML), nous constatons que le développement du microbiome intestinal, les capacités cognitives des enfants et la structure cérébrale sont intimement liés, les taxons microbiens et les fonctions géniques étant capables de prédire la performance cognitive et la structure cérébrale.
Axe intestin/ cerveau : Nos intestins sont tapissés de bactéries, de virus et de champignons. Le microbiote qui affecte le métabolisme indiquent que les bactéries intestinales seraient impliquées dans les comportements.
RÉSULTATS
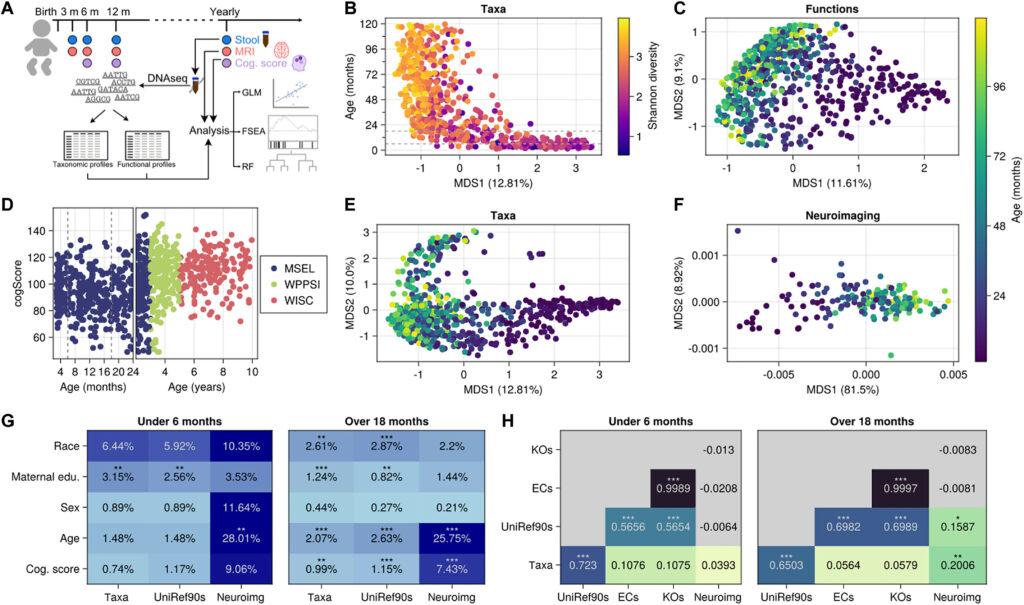
La cohorte RESONANCE fait partie d’une étude sur le développement de l’enfant
Nous avons étudié la co-développement du cerveau et du microbiome chez 381 enfants en bonne santé et se développant de manière neurotypique (172 filles) entre 40 jours et 10 ans en utilisant diverses mesures microbiennes et neurocognitives orthogonales. Celles-ci comprenaient le séquençage métagénomique shotgun, des évaluations cognitives et comportementales appropriées à l’âge en utilisant les scores composites à l’échelle complète de MSEL, WPPSI-4 et WISC-V, et des mesures neuroimaging de la morphologie corticale et sous-corticale. Différents instruments d’évaluation ont été utilisés en fonction de l’âge de l’enfant en raison de la plage d’âge unique et des propriétés psychométriques mesurées par chacun (c’est-à-dire, MSEL pour les enfants de moins de 4 ans, WPPSI-3 pour les 3 à 6 ans, et WISC-V pour les enfants de plus de 6 ans), mais ont été normalisés à une échelle commune. Comme prévu, les plus grandes différences dans les taxons microbiens ont été observées en fonction de l’âge des enfants, les enfants plus âgés étant principalement stratifiés en taxons dominants Bactéroïdètes, Firmicutes ou Prevotella copri à haute abondance. La variation globale d’un sujet à l’autre dans les gènes microbiens intestinaux et les profils de volume cérébral était également largement influencée par l’âge des sujets.
Comme l’ont démontré plusieurs études antérieures, des associations entre des taxons spécifiques du microbiote intestinal et une neurocognition atypique ainsi que des troubles neurologiques ont été établies. Nous avons donc cherché à déterminer si des taxons spécifiques ou des fonctions géniques étaient associés au développement cognitif normal chez les enfants. Pour tester si des variations dans les taxons microbiens intestinaux, leurs gènes ou leur métabolisme étaient associées au développement neurocognitif, nous avons utilisé une analyse de variance par permutation (PERMANOVA) incluant la diversité β des taxons microbiens et des fonctions géniques, les mesures de volume dérivées de l’imagerie cérébrale des structures corticales et sous-corticales, et l’aptitude cognitive générale. Étant donné le changement écologique majeur dans le microbiome qui se produit avec l’introduction d’aliments solides, et la plage relativement large d’âges pendant lesquels les nourrissons passent du lait aux aliments solides, nous avons séparément considéré les âges généralement pré-transition (avant 6 mois) et ceux généralement post-transition (plus de 18 mois). Cependant, même avec cette catégorisation, nous observons une variation croissante dans les microbiomes intestinaux des enfants de plus de 18 mois. Nous avons également constaté que la variation globale des espèces microbiennes chez les enfants de plus de 18 mois était significativement associée à la variation du score de la fonction cognitive (R2 = 0,0099, q < 0,01), tout comme la variation dans les fonctions géniques microbiennes (R2 = 0,0115, q < 0,001). La variation dans les taxons microbiens et les gènes n’était pas significativement associée à la fonction cognitive chez les enfants de moins de 6 mois, bien que cela puisse être dû à la faible diversité taxonomique et à l’absence générale de chevauchement entre les taxons chez les nourrissons. Comme prévu, l’âge était significativement associé à la diversité bêta microbienne [taxons R2 = 0,0207, et fonctions géniques annotées avec des clusters UniRef90 de 90% de similarité (28) : R2 = 0,0258, q < 0,001] et fortement associé à la variation dans les profils d’imagerie neurologique (IRM) (R2 = 0,258, q < 0,001).
Conformément à des études antérieures, différents types de mesures microbiennes capturaient une variation chevauchante, les profils d’espèces et les profils de fonctions géniques, tous deux générés à partir de séquences métagénomiques, étant étroitement liés (P < 0,001). D’autres regroupements fonctionnels [niveau 4 de la classification de la Commission des enzymes (ECs) et orthologues de l’Encyclopédie des gènes et génomes de Kyoto (KOs)] présentaient seulement une légère chevauchement avec les profils taxonomiques dans les deux cohortes d’âge, malgré qu’ils soient dérivés d’étiquettes UniRef90. Chez les enfants de plus de 18 mois, une certaine variation (taxons : 20,1%, P < 0,01 ; fonctions géniques : 15,9%, P < 0,05) dans l’imagerie neurologique chevauchait avec les mesures microbiennes, bien que cela puisse être dû à la variation résiduelle liée à l’âge dans les deux mesures.
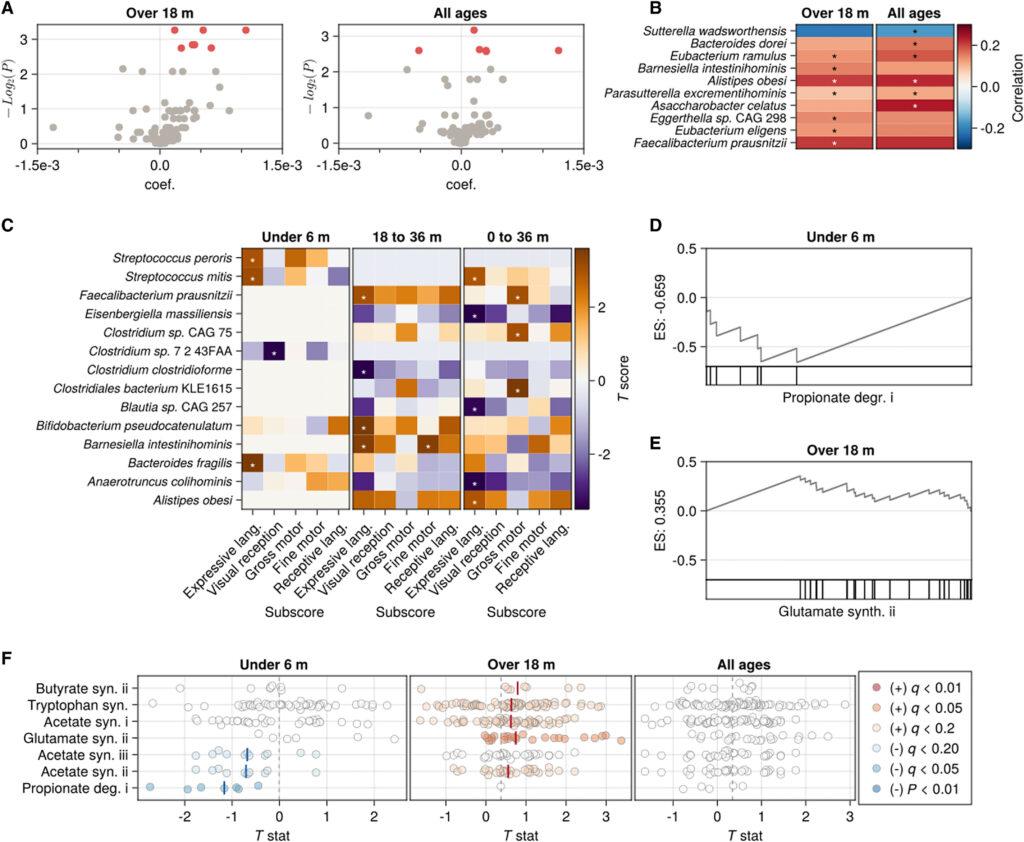
Des espèces microbiennes et des gènes neuroactifs sont associés à la performance cognitive Pour évaluer si des espèces microbiennes individuelles étaient associées à la fonction cognitive, nous avons ajusté des modèles de régression linéaire multivariables (LMs) pour l’abondance relative de chaque espèce ayant une prévalence d’au moins 15% dans un groupe d’âge donné (N-espèces = 116 de 0 à 120 mois, N-espèces = 54 de 0 à 6 mois, N-espèces = 136 de 18 à 120 mois). Aucune espèce n’était significativement associée (q < 0,20) à la fonction cognitive chez les enfants de moins de 6 mois après ajustement pour l’âge et l’éducation maternelle. En revanche, chez les enfants de plus de 18 mois, plusieurs espèces microbiennes étaient significativement enrichies chez les enfants ayant des scores de fonction cognitive plus élevés, dont Alistipes obesi, Asaccharobacter celatus [également connue sous le nom d’Adlercreutzia equolifaciens subsp. celatus (32)], et des espèces probiotiques productrices d’AGCC telles que Eubacterium eligens et Faecalibacterium prausnitzii. Sutterella wadsworthensis était la seule espèce microbiennne significativement associée négativement au score de la fonction cognitive (q = 0,165).
(fig 1) – Micrographie électronique à basse température d’un groupe de bactéries E. coli. (fig 2) – Le corps humain accueille autour de 100 000 milliards de microbes. (fig 3) – Microbiote : des bactéries ont été trouvées dans le cerveau.
Les évaluations cognitives (MSEL, WPPSI, WISC) sont des composites de plusieurs sous-échelles évaluant différents domaines cognitifs.
Bien que les scores composites soient largement analogues entre les âges et les modalités de test, les sous-échelles évaluées dans chaque test sont différentes. Pour évaluer si les taxons microbiens étaient associés au développement de domaines cognitifs spécifiques, nous avons ajusté des modèles linéaires multivariés pour chaque espèce présente dans au moins 10 % des échantillons d’enfants de moins de 6 mois, de 18 à 36 mois, ou de tous les âges avec un score MSEL simultané. Plusieurs taxons associés aux scores composites étaient également associés à un ou plusieurs sous-scores, notamment F. prausnitzii et A. obesi. Bien que aucun taxon ne soit significativement associé aux scores composites chez les enfants de moins de 6 mois, deux espèces du genre Streptococcus (S. peroris et S. mitis), ainsi que B. fragilis, étaient positivement associées au langage expressif dans ce groupe d’âge. S. mitis était également associé au langage expressif en considérant tous les âges, tandis qu’Eisenbergiella massiliensis, Anaerotruncus colihominis, et une espèce de Blautia non classée étaient négativement associées.
Étant donné que différentes espèces microbiennes peuvent occuper la même niche métabolique chez différents individus, nous avons émis l’hypothèse que les gènes microbiens regroupés par activité fonctionnelle seraient associés à la cognition. Pour tester cela, nous avons effectué une analyse d’enrichissement de l’ensemble des caractéristiques (FSEA) sur des groupes de gènes ayant un potentiel neuroactif et un score de capacité cognitive simultané et avons constaté que plusieurs voies métaboliques étaient soit significativement enrichies, soit appauvries chez les enfants ayant des scores de capacité cognitive plus élevés. Par exemple, les gènes de dégradation des acides gras à courte chaîne (AGCC) à 3 carbones, comme le propionate, étaient significativement appauvris chez les enfants de moins de 6 mois ayant des scores de capacité cognitive plus élevés [Tableau 2 ; dégradation du propionate I, score d’enrichissement (ES) = −0,66, valeur corrigée (q) = 0,029], de même que les gènes de synthèse de l’AGCC à 2 carbones acétate (synthèse d’acétate II, ES = −0,491, q = 0,059 ; synthèse d’acétate III, ES = −0,409, q = 0,133). Chez les enfants de plus de 18 mois, ces associations métaboliques étaient inversées, avec à la fois la synthèse d’acétate et la dégradation du propionate enrichies chez les enfants ayant des scores cognitifs plus élevés (synthèse d’acétate I, ES = 0,192, q = 0,145 ; synthèse d’acétate II, ES = 0,248, q = 0,15 ; dégradation du propionate I, ES = 0,154, q = 0,178), de même que la synthèse de l’AGCC à 4 carbones, le butyrate (synthèse de butyrate II, ES = 0,414, q = 0,178). La synthèse de l’acide aminé glutamate était également significativement enrichie chez les enfants ayant des scores de fonction cognitive plus élevés dans ce groupe d’âge (synthèse de glutamate II, ES = 0,355, q = 0,023). Ensemble, ces résultats suggèrent que l’activité métabolique des micro-organismes, en particulier le métabolisme (synthèse et dégradation) de composés neuroactifs, peut avoir des effets sur le développement cognitif.
Les profils taxonomiques et fonctionnels du microbiote intestinal prédisent la fonction cognitive mesurée simultanément.
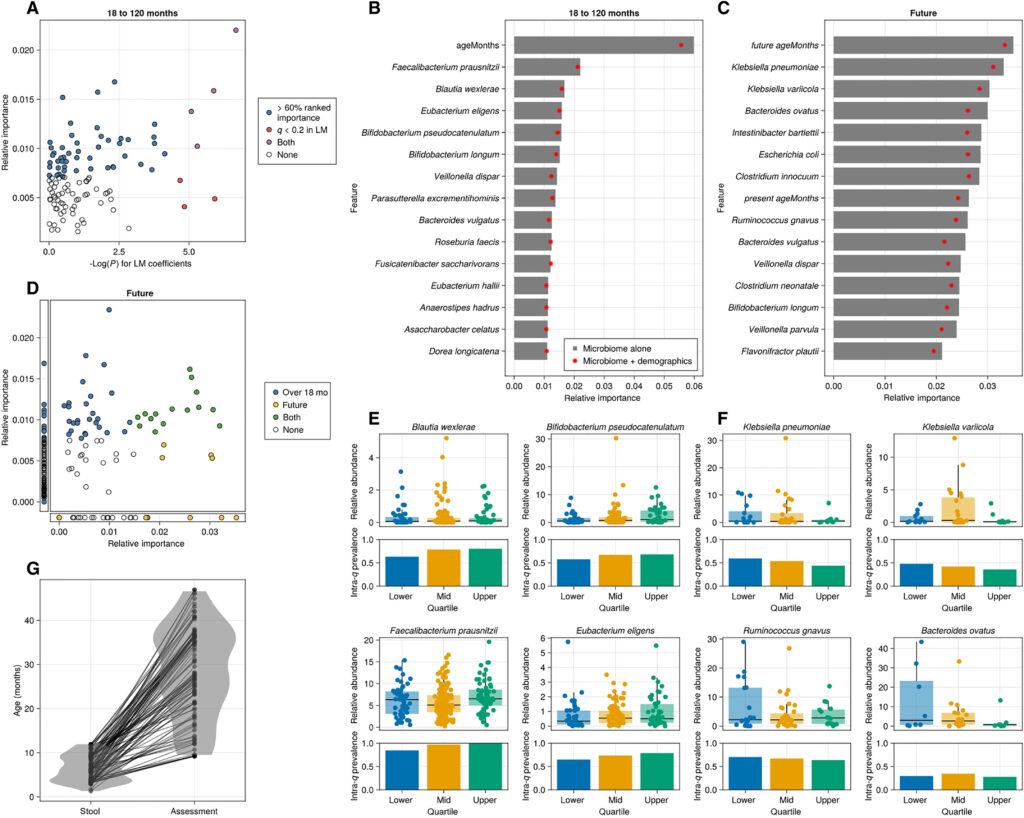
L’analyse d’enrichissement de l’ensemble des caractéristiques (FSEA) repose sur la compréhension des relations fonctionnelles entre les gènes individuels. Cependant, étant donné que les relations entre les taxons individuels sont largement inconnues, nous nous sommes tournés vers les modèles de forêts aléatoires (RF), une famille de méthodes d’apprentissage automatique non supervisées et non paramétriques qui permet d’identifier des motifs sous-jacents dans un grand nombre de caractéristiques individuelles (ici, les espèces microbiennes). Des études antérieures ont rapporté l’utilisation réussie des RF pour traiter des données hautement dimensionnelles et clairsemées dans le domaine de la génomique, ainsi que dans d’autres travaux où il a été utilisé pour prédire des conditions cognitives liées à la maladie d’Alzheimer dans différents scénarios. Étant donné que les profils microbiens intestinaux, ainsi que le développement neurocognitif, peuvent partiellement refléter des facteurs socio-économiques et démographiques, nous avons évalué les performances des régresseurs RF où l’éducation maternelle [utilisée ici comme un indicateur du statut socio-économique (SES)], le sexe et l’âge étaient inclus en tant que prédicteurs possibles, soit individuellement, soit en combinaison avec les profils taxonomiques microbiens.
Tout comme les modèles linéaires (LM), les modèles de forêts aléatoires (RF) pour les enfants de moins de 6 mois ont donné de mauvais résultats (corrélation moyenne sur l’ensemble de test -0,13, erreur quadratique moyenne des racines (RMSE) moyenne 13,01), mais les RF étaient constamment capables d’apprendre la relation entre les taxons et les scores de fonction cognitive chez les enfants de plus de 18 mois (corrélation moyenne sur l’ensemble de test 0,429, RMSE moyen 18,25). Les espèces importantes dans les RF chevauchaient celles qui étaient significatives lors du test de la relation avec les LM. Les deux analyses ont identifié F. prausnitzii, E. eligens, Parasutterella excrementihominis et A. obesi comme importantes (q < 0,2 pour les LM, importance cumulée <60 % pour les RF). D’autres taxons significatifs dans les LM n’ont pas été identifiés par les modèles RF, notamment Eggerthella sp. CAG 298 et Barnesiella intestinihominis. Eubacterium ramulus était juste en dehors de notre seuil de coupure (importance cumulée de 64 %). Les modèles RF ont également identifié 48 espèces supplémentaires importantes pour discriminer les scores de fonction cognitive. L’âge du sujet était constamment classé en haute importance de fonction, ce qui pourrait indiquer que les branches de décision basées sur les taxons microbiens ont une pureté accrue lorsqu’on considère l’âge du sujet ou que l’âge lui-même est un prédicteur utile.
Les profils microbiens intestinaux au cours de la première année de vie prédisent la fonction cognitive future.
Bien que les modèles linéaires (LM) n’aient identifié aucun taxon dans la tranche d’âge de 0 à 6 mois qui était associé à la fonction cognitive mesurée simultanément, nous avons considéré que les trajectoires de développement débutant in utero pourraient ne pas être immédiatement modifiées par les expositions microbiennes, de sorte que les influences microbiennes précoces dans la vie pourraient prendre du temps pour se manifester. Pour tester cette hypothèse, nous avons examiné si les modèles de forêts aléatoires (RF) formés sur les profils taxonomiques au cours de la première année de vie pouvaient prédire les performances cognitives futures mesurées au moins 6 mois après le prélèvement de l’échantillon de selles. Les modèles ont pu de manière cohérente apprendre les espèces microbiennes associées aux performances cognitives lors d’une visite ultérieure (moyenne du temps entre les visites : 21,5 mois, écart type = 9,9 mois, corrélation moyenne sur l’ensemble de test : 0,27).
/// La cohorte RESONANCE est une cohorte diversifiée avec un échantillonnage longitudinal accéléré ///
Presque tous les taxons importants dans les modèles de forêts aléatoires (RF) qui prédisaient la fonction cognitive chez les enfants de plus de 18 mois étaient également importants dans les modèles qui prédisaient la fonction cognitive future, s’ils étaient présents. En même temps, de nombreuses espèces supplémentaires étaient importantes pour les modèles de prédiction future qui n’étaient pas importantes dans les modèles mesurant la fonction cognitive simultanée. En particulier, l’importance des caractéristiques pour les modèles de prédiction future était dominée par deux espèces de Klebsiella, K. pneumoniae et K. varicola. Beaucoup des autres espèces qui figuraient en tête de l’importance des caractéristiques étaient également des pathogènes opportunistes, ou avaient déjà été signalées comme liées à des maladies (par exemple, Ruminococcus gnavus, Escherichia coli).
Les taxons microbiens intestinaux sont associés à des fonctions cognitives spécifiques.
Étant donné que le développement de différents domaines de la fonction cognitive (par exemple, le développement moteur brut par rapport à la perception visuelle) peut se produire à différents moments et être affecté par différents signaux moléculaires, nous avons cherché à déterminer si les associations des taxons microbiens avec la fonction cognitive sont générales pour tous les sous-domaines cognitifs. Pour ce faire, nous avons évalué les sous-échelles individuelles du MSEL. Les profils taxonomiques associés aux données démographiques ont montré un potentiel prédictif pour trois des cinq sous-échelles examinées, à savoir le langage expressif (RMSE de l’ensemble de test 12,05, corrélation de l’ensemble de test 0,368), le développement moteur brut (RMSE de l’ensemble de test 11,07, corrélation de l’ensemble de test 0,137), et la réception visuelle (RMSE de l’ensemble de test 15,2, corrélation de l’ensemble de test 0,333). Entre les sous-échelles, tant la prédictibilité (mesurée par les indicateurs de l’ensemble de test) que l’importance relative de plusieurs taxons sont variables. Bien que certaines espèces telles que B. longum et F. prausnitzii aient une importance relative similaire dans toutes les sous-échelles, d’autres sont sensiblement différentes. Notamment, Bifidobacterium pseudocatenulatum, Blautia wexlerae et E. eligens sont nettement plus importants pour la prédiction du langage expressif, tandis que Roseburia faecis, Streptococcus salivarius et Fusicatenibacter saccharivorans sont distinctement prédictifs du développement moteur brut, et Clostridium innocuum et Bacteroides vulgatus se distinguent dans les modèles ciblant la réception visuelle.
Les profils taxonomiques microbiens intestinaux prédisent les différences de structure cérébrale.
Étant donné que des microbes individuels pourraient être associés à des domaines particuliers de la fonction cognitive et pas à d’autres, s’il existe des effets causaux du métabolisme microbien sur la fonction cognitive, ils pourraient se refléter dans des changements de la neuroanatomie. Nous avons de nouveau utilisé la modélisation de forêts aléatoires (RF) pour associer les profils taxonomiques intestinaux avec les régions cérébrales individuelles identifiées dans les IRM (enfants de plus de 18 mois, N = 124). Certaines régions cérébrales ont été plus facilement prédites par les modèles RF formés sur les taxons microbiens (Tableaux 5 et 6 et tableau S6), en particulier celles qui étaient fortement corrélées à l’âge. Cela incluait le gyrus lingual gauche/droit (corrélation RF moyenne, gauche = 0,444, droit = 0,441 ; importance pour l’âge, gauche = 6,8 %, droit = 7,7 %) et le cortex péricalcarine gauche/droit (corrélation RF moyenne, gauche = 0,267, droit = 0,315 ; importance pour l’âge, gauche = 3,6 %, droit = 6,2 %). Dans de nombreux cas, cependant, l’âge n’était pas une variable importante dans les modèles performants, comme celui de la région accumbens gauche (corrélations RF moyennes = 0,275 ; importances relatives de l’âge = 1,1 %).
Alors que les modèles pour de nombreuses régions étaient fortement concordants entre les hémisphères gauche et droit en termes de performances du modèle et d’importance des caractéristiques microbiennes, d’autres régions présentaient des différences bilatérales substantielles. Par exemple, la zone accumbens gauche présente l’une des corrélations les plus élevées avec l’ensemble de test de nos modèles de région cérébrale (R = 0,288), comparée à la droite, qui présentait une corrélation négligeable avec l’ensemble de test (R = 0,005). L’importance des caractéristiques pour les modèles de la zone accumbens gauche était dominée par trois espèces de Bacteroides, B. vulgatus (importance de 3,3 %), B. ovatus (importance de 3,2 %) et B. uniformis (importance de 2,8 %).
Bien que les modèles RF pour plusieurs régions cérébrales aient de nombreux taxons microbiens importants, d’autres étaient dominés par un petit nombre de taxons, dont beaucoup ont également été identifiés dans les LM et les modèles RF de la fonction cognitive. En général, pour toutes les segments, un nombre constant d’espèces compris entre 28 et 42 (médiane = 38, moins de 30 % des caractéristiques d’entrée) étaient responsables de la moitié de l’importance cumulative, quel que soit la performance du modèle. Nous avons observé deux principaux schémas de distribution de l’importance des espèces sur les segments cérébraux ; certaines espèces ont montré des contributions élevées à plusieurs segments différents, tandis que d’autres contribuaient modestement à un ou deux segments cérébraux. Des cas notables du premier schéma comprenaient neuf espèces : Anaerostipes hadrus, B. vulgatus, B. ovatus, F. saccharivorans, Ruminococcus torques, Eubacterium rectale, Coprococcus comes, A. celatus, et B. wexlerae, qui, combinées, représentent environ 10 % des importances relatives cumulatives, calculées après pondération de la pertinence sur les segments cérébraux rapportés.
Parmi celles-ci, A. hadrus est la variable avec la plus haute importance relative moyenne sur tous les modèles rapportés (moyenne = 1,4 % ; min = 0,8 % ; max = 3,6 %). B. wexlerae, qui était importante dans les modèles RF de la fonction cognitive, était également importante dans les modèles des zones entorhinales, fusiformes et linguales et avait l’importance la plus élevée dans les modèles prédisant la taille du parahippocampe gauche et droit. Un autre groupe d’importance de modèle contenait des taxons associés au cerveau basal, en particulier le cingulaire et l’accumbens. Ce groupe comprend B. ovatus et B. uniformis, mais aussi Alistipes finegoldii et S. salivarius.
A. celatus, une espèce productrice d’équol, est un exemple du deuxième schéma de contribution, où un taxon ne contribue significativement qu’à un très petit nombre de segments. Cette espèce était très importante dans la prédiction du volume relatif du cingulaire antérieur droit (importance relative de 2,8 %). Un autre exemple de ce schéma est Coprococcus eutactus, fortement associé à la zone précentrale gauche (importance de 3,5 %). Enfin, C. comes a une distribution d’importance qui se divise presque également entre les deux clusters précédemment rapportés. Sa contribution à la prédiction du cingulaire postérieur gauche est la plus élevée pour un microbe dans les modèles RF (importance de 3,7 %) ; seul l’âge avait une importance relative plus élevée dans n’importe quel modèle. En même temps, C. comes est l’un des prédicteurs les plus importants pour les zones voisines du cingulaire postérieur, telles que la pars opercularis (importances, gauche = 1,9 %, droite = 2,5 %), ainsi que les régions supérieures comme la zone précentrale gauche (importance de 2,0 %) et les lobes paracentraux (importances, gauche = 2,0 %, droite = 1,5 %).
/// Les taxa et les groupes fonctionnels de gènes sont associés à la fonction cognitive ///
Débat
La relation entre le microbiome intestinal et la fonction cérébrale via l’axe intestin-cerveau-microbiome a été de plus en plus acceptée, principalement en raison d’études épidémiologiques humaines examinant la neurocognition atypique [par exemple, anxiété et dépression, neurodégénérescence, trouble déficitaire de l’attention/hyperactivité et autisme] et d’études mécanistiques sur des modèles animaux. Les résultats de ces études suggèrent la possibilité que les microbes intestinaux et leur métabolisme puissent être causalement impliqués dans le développement cognitif, mais cette étude est, à notre connaissance, la première à enquêter directement sur les espèces microbiennes et leurs gènes en relation avec le développement typique chez les jeunes enfants en bonne santé. Comprendre l’axe intestin-cerveau-microbiome tôt dans la vie est particulièrement important, car les différences ou interventions précoces peuvent avoir des conséquences démesurées et à plus long terme que celles à un âge plus avancé en raison de la nature dynamique et plastique du microbiome intestinal et du cerveau. De plus, même en l’absence d’impacts causaux du métabolisme microbien, identifier des facteurs de risque pouvant indiquer d’autres interventions précoces aurait également de la valeur.
Nous avons identifié plusieurs espèces de la famille Eggerthelaceae associées à la fonction cognitive, notamment A. celatus [une sous-espèce d’A. equolofaciens] et Eggerthella lenta. De nombreux membres de cette famille sont connus en partie en raison de leurs activités métaboliques uniques. Par exemple, A. equolofaciens produit l’estrogène non stéroïdien équol à partir d’isoflavènes présents dans le soja. Cela est particulièrement intrigant, car A. celatus était également la caractéristique la plus importante dans les modèles RF du cingulaire antérieur droit, qui a été lié à la cognition sociale et à la prise de décision basée sur la récompense, et l’estrogène a déjà montré une activité dans cette région cérébrale. Un autre Eggerthelaceae, Gordonibacter pamelaeae, a également un métabolisme unique qui peut affecter le cerveau, car il peut métaboliser l’acide ellagique (trouvé dans les grenades et certaines baies) en urolithine, qui a été démontré dans certaines études avoir un effet neuroprotecteur. Bien que G. pamelaeae n’ait pas été identifié comme une caractéristique importante dans les modèles RF, il était également relativement peu fréquent dans notre ensemble de données, se produisant dans seulement 5 % des échantillons à une abondance supérieure à 2 % (contrairement à A. celatus, qui était dans 32 % des échantillons). Bien que nous n’ayons pas trouvé de preuves publiées de rôles neurologiques pour E. lenta, elle a été largement étudiée pour sa capacité à métaboliser des composés médicamenteux tels que la digoxine. La polyvalence métabolique de ce clade et le grand nombre d’espèces associées à la cognition font de ces microbes des cibles idéales pour des études mécanistiques approfondies.
Nous n’avons identifié aucune association statistique entre les espèces individuelles et la performance cognitive dans notre plus jeune sous-ensemble (0 à 6 mois). Nous pensons que cela peut être dû au fait que les trajectoires du développement cérébral commencent in utero et que les influences microbiennes (qui ne peuvent commencer qu’à la naissance) mettent du temps supplémentaire à se manifester. En accord avec cette hypothèse, les modèles RF ont pu apprendre des associations entre les microbiomes de la petite enfance et les scores de la fonction cognitive future. Les espèces les mieux classées pour ces modèles comprenaient plusieurs taxons importants pour prédire les scores d’évaluation cognitive simultanée après 18 mois, notamment des pathobiontes de la famille Enterobacteriaceae tels qu’Escherichia et Klebsiella. Les représentants de ces genres sont connus pour être des colonisateurs précoces et à long terme persistants de l’intestin des nourrissons qui peuvent, dans certaines situations, favoriser une réponse inflammatoire. En revanche, plusieurs représentants réduisant l’inflammation du genre Bifidobacterium, du métaboliseur précoce des oligosaccharides du lait humain B. longum au métaboliseur tardif des glucides complexes B. pseudocatenulatum, associés couramment au sevrage à la nourriture solide à un jeune âge, ont également montré une importance relative élevée. L’implication de la plupart des taxons importants dans différents aspects de la réponse inflammatoire résonne avec des résultats antérieurs qui associent l’inflammation chronique à des changements dans les réponses neuronales à certains types de stimuli.
D’autres bactéries associées à la fois à la capacité cognitive et à la structure cérébrale ont également des connexions possibles avec des études antérieures. Par exemple, les modèles RF prédisant la taille du noyau accumbens gauche étaient dominés par les espèces de Bacteroides, en particulier B. vulgatus, B. uniformis et B. ovatus. Il a été démontré que B. ovatus produit plusieurs composés neuroactifs, notamment des acides gras à chaîne courte (AGCC) et des neurotransmetteurs tels que l’acide γ-aminobutyrique (GABA). Une autre espèce de Bacteroides, B. fragilis, a été capable d’améliorer les symptômes similaires à ceux du trouble du spectre autistique (TSA) dans un modèle murin en consommant un métabolite dépendant des microbes, le 4-éthylphénylsulfate (4EPS), tandis que B. ovatus est un producteur primaire d’un précurseur du 4EPS et a augmenté les comportements anxieux chez les souris. Le fait que l’importance de ces espèces dans les modèles RF et la haute performance des modèles RF dans la prédiction de cette région étaient unilatérales (affectant la zone accumbens gauche mais pas la droite) était également notable. La plupart des individus en bonne santé ont une asymétrie droite dans le noyau accumbens, qui est impliqué dans l’apprentissage de la récompense et les comportements de prise de risque. Une asymétrie réduite entre le noyau accumbens droit et gauche a été associée à un trouble de l’utilisation de substances chez les jeunes adultes. La zone accumbens est également associée au contrôle de la récompense, et chez les personnes diagnostiquées avec un trouble du déficit de l’attention avec hyperactivité (TDAH), il a été démontré qu’elle présente une neuromorphologie divergente.
Notre capacité à sonder les sous-échelles cognitives individuelles a fourni des résultats qui correspondent également à des résultats antérieurs et fournissent un aperçu des connexions potentielles entre la cognition et la neuroanatomie intestino-microbienne. Par exemple, une étude longitudinale sur le développement neurocognitif a mis en évidence des associations bilatérales entre le thalamus et la composante du langage expressif des échelles Mullen. En accord avec cette découverte, nous avons constaté que des taxons tels que B. wexlerae et A. hadrus figuraient parmi les prédicteurs les plus importants des deux côtés du thalamus et de la sous-échelle du langage expressif. Un autre exemple de ces résultats est une association précédemment rapportée entre le cortex opérculaire central (COC) et le développement moteur global. Dans ce cas, nous avons identifié que F. saccharivorans, producteur de butyrate, est parmi les 15 % de prédicteurs les plus importants pour la sous-échelle du développement moteur global et trois composants bilatéraux du COC (pars orbitalis gauche et droite, pars opercularis et pars triangularis). Les résultats antérieurs sont également liés aux relations entre le noyau caudé et le cervelet, et le score de réception visuelle. S. salivarius a été constamment trouvé parmi les 20 % de prédicteurs supérieurs pour cette sous-échelle, ainsi que des deux côtés du thalamus et de la substance blanche du cervelet, ainsi que des lobules I à X du vermis cérébelleux.
Comme le montre clairement le rôle potentiellement opposé de B. fragilis et de B. ovatus, les espèces individuelles d’un même genre peuvent jouer des rôles différents. Ainsi, l’utilisation du séquençage métagénomique à shotgun peut être cruciale pour identifier les influences microbiennes sur le cerveau, car cela permet une résolution au niveau des espèces des taxons microbiens. Une étude antérieure sur la cognition chez des sujets de 3 ans utilisant le séquençage d’amplicons du gène 16S de l’ARNr a montré que les genres de la famille Lachnospiraceae ainsi que les Clostridiales non classés (maintenant Eubacteriales) étaient associés à des scores plus élevés sur le questionnaire Ages and Stages. Cependant, chacune de ces clades englobe des dizaines de genres aux fonctions diverses, qui peuvent avoir des effets différents. Plusieurs des taxons positivement associés à la fonction cognitive dans cette étude, tels que B. wexlerae, Dorea longicatena, R. faecis et A. finegoldii, appartiennent aux Clostridiales, tout comme R. gnavus, que nous avons trouvé être négativement associé à la fonction cognitive. Ce niveau de résolution au niveau des espèces n’est généralement pas possible avec le séquençage d’amplicons.
En plus d’une meilleure résolution au niveau des espèces, le séquençage métagénomique à shotgun permet également d’obtenir des informations fonctionnelles sur les gènes. Nous avons montré que les gènes impliqués dans le métabolisme des SCFA, tant dans leur dégradation que dans leur synthèse, sont associés aux scores de la fonction cognitive. Les SCFA sont produits par la fermentation anaérobie des fibres alimentaires et ont été associés à la régulation du système immunitaire ainsi qu’au fonctionnement direct du cerveau. Le métabolisme microbien d’autres molécules potentiellement neuroactives était également associé à la capacité cognitive. Par exemple, nous étions particulièrement intéressés par les gènes impliqués dans le métabolisme du glutamate, un neurotransmetteur critique qui contrôle la signalisation neuronale excitatrice/inhibitrice, ainsi que du GABA. L’équilibre entre ces deux molécules dans le cerveau contrôle la plasticité neuronale et l’apprentissage, en particulier dans le cerveau en développement. Cependant, bien que l’abondance différentielle des gènes impliqués dans le métabolisme de composés neuroactifs comme ceux-ci soit suggestive, il est difficile de raisonner sur la relation entre les niveaux de ces gènes et les concentrations dans le côlon des molécules sur lesquelles agissent les enzymes produites par ces gènes, car la présence ou l’absence de métabolites peuvent sélectionner des microbes ayant la capacité de dégrader ou de produire ce métabolite, respectivement. Pour cette raison, des études futures couplant le séquençage métagénomique à shotgun avec la métabolomique des selles pourraient améliorer notre compréhension de la relation entre le métabolisme microbien et le développement cognitif. L’analyse métabolomique sérique pourrait être encore plus informative, car les métabolites mesurés dans les selles ne sont pas nécessairement indicatifs de l’exposition, mais le prélèvement de sang suffisant chez les enfants, en particulier les très jeunes enfants, est difficile. De plus, une analyse au niveau de la souche liant le contenu génique spécifique des espèces d’intérêt pourrait affiner davantage les efforts ciblés visant à identifier des signatures métaboliques spécifiques des interactions microbe-cerveau.
L’utilisation de plusieurs évaluations cognitives appropriées à l’âge et pouvant être normalisées à une échelle commune nous a permis d’analyser les associations microbiennes sur plusieurs périodes de développement, mais cela comporte plusieurs inconvénients. En particulier, la fiabilité des tests, ainsi que les différences systématiques entre les administrateurs de tests, peuvent introduire un bruit substantiel dans ces observations, en particulier chez les enfants les plus jeunes. De plus, notre période d’étude a chevauché le début de la pandémie de la maladie à coronavirus 2019 (COVID-19), et nous, ainsi que d’autres, avons observé une certaine réduction des scores mesurés pour les enfants évalués après la mise en œuvre des mesures de confinement. Dans notre ensemble de sujets pour cette étude, ces effets sont plus prononcés dans certains groupes d’âge en raison de la période où le recrutement actif pour l’étude a eu lieu.
Cette analyse nous a permis d’établir des liens entre les taxons microbiens et leur potentiel fonctionnel avec la cognition et la structure cérébrale. Bien que nous n’ayons pas testé directement les relations causales entre les taxons microbiens intestinaux et leurs gènes, l’intestin et le cerveau, cette étude fournit des associations claires et statistiquement significatives qui pourraient servir de cibles pour les efforts futurs dans des modèles précliniques. Par exemple, des modèles murins de développement cérébral, d’apprentissage et de fonctions exécutives pourraient être évalués chez des animaux sans germes avec l’ajout de microbes caractérisés et manipulés métaboliquement. Les futures études devraient également se concentrer sur la caractérisation du microbiome et du développement neurocognitif au cours de la petite enfance dans différentes régions géographiques et modes de vie, en couvrant des populations traditionnellement sous-étudiées, telles que les communautés urbaines à faibles ressources, péri-urbaines et rurales, pour obtenir une compréhension plus complète de la variabilité au sein des différents microbiomes intestinaux reflétés sur la neurocognition. Ces études nous fourniraient également une richesse de données sur différentes souches de la même espèce pour mieux comprendre l’effet des gènes et de leurs produits. De plus, des études de culture et d’enrichissement de la communauté microbienne combinées à la manipulation génétique et aux approches génomiques pour comprendre le métabolisme microbien au niveau moléculaire sont essentielles, car les fonctions métaboliques façonnent et influencent l’hôte humain et sa santé. La découverte de métabolites neuroactifs pourrait nous fournir des biomarqueurs pour la détection précoce ou des molécules médicinalement utiles qui pourraient être appliquées dans des interventions.
/// Les micro-organismes résidant dans l’intestin et leurs gènes sont associés à la cognition et à la neuroanatomie chez les enfants ///

Alessandro Lettiere
Annonces PARTENAIRES



